

Excavata
| Excavata | |||||
|---|---|---|---|---|---|
 | |||||
| Biyolojik sınıflandırma (parafiletik olarak geçerliliğini yitirmiş) | |||||
| |||||
| Filum ve sınıflar | |||||

Excavata, Ökaryot üst alemine ait eski bir tek hücreli organizma üst grubudur.[1][2][3] İlk olarak 1999'da Simpson ve Patterson tarafından önerilmiş [4][5] ve 2002'de Thomas Cavalier-Smith tarafından resmi bir takson olarak tanıtılmıştır. Çeşitli serbest yaşayan ve simbiyotik formlar içerir ve Giardia ve Trichomonas gibi bazı önemli insan parazitlerini de içinde barındırır.[6] Daha önce artık modası geçmiş Protista krallığına dahil olduğu düşünülüyordu.[7] Kamçılı yapılarına göre sınıflandırıldılar [5] ve en temel kamçılı soy olarak kabul edildiler.[8]
Filogenomik analizler, Excavata'nın polifiletik olduğunu ve tek bir doğal grup oluşturmadığını gösterir.[9] Bu nedenle Excavata, hepsi yakından ilişkili olmayan üç gruba ayrılmıştır: Discobid, Metamonad ve Malawimonad.[10][11][12] Euglenozoa dışında hepsi fotosentetik değildir.
Özellikler
[değiştir | kaynağı değiştir]Çoğu excavate tek hücreli, heterotrofik flagellatlardır. Sadece Euglenozoa fotosentetiktir. Bazılarında (özellikle anaerobik bağırsak parazitlerinde), mitokondri büyük ölçüde azalmıştır.[6] Bazılarında "klasik" mitokondri yoktur ve "amitokondriat" olarak adlandırılırlar ancak çoğu büyük ölçüde değiştirilmiş formda (örneğin bir hidrojenozom veya mitozom) bir mitokondriyal organel içerir. Mitokondrili olanlar arasında mitokondriyal cristae ; tübüler, diskoidal veya bazı durumlarda laminer olabilir. Çoğunda iki, dört veya daha fazla flagella bulunur.[5] Birçoğunun, mikrotübüller tarafından desteklenen, karakteristik bir ince yapıya sahip göze çarpan bir ventral besleme oluğu vardır. Organizmalara adını veren bu oluğun "kazılmış" görünümüdür.[4][7] Bununla birlikte bu özelliklere sahip olmayan çeşitli gruplar, genetik kanıtlara (öncelikle moleküler dizilerin filogenetik ağaçları) dayalı olarak excavate olarak kabul edilebilir.[7]
Acrasidae balçık küfleri, sınırlı çok hücrelilik sergileyen tek excavatedir. Diğer hücresel cıvık mantarlar gibi, hayatlarının çoğunu tek hücreler olarak yaşarlar ancak bazen daha büyük kümeler halinde birleşirler.
Önerilen grup
[değiştir | kaynağı değiştir]Excavate ilişkileri her zaman belirsizdi, bu da onların monofiletik bir grup olmadıklarını gösteriyor.[13] Malawimonad'lar genellikle diğer Excavata ile dallanmaz.[14]
Excavatelerin birden çok grubu içerdiği düşünülüyordu:
| Krallık/Süperfilum | Dahil olan taksonlar | Temsili cins (örnekler) | Tanım |
|---|---|---|---|
| Diskoba veya JEH veya Eozoa | Tsukubea | Tsukubamonas | |
| Öglenozoa | Euglena,tripanosoma | Birçok önemli parazit, plastidli büyük bir grup (kloroplastlar) | |
| heterolobosea (Perkolozoa) | Naegleria, Acrasis | Çoğu kamçılı ve amoeboid formlar arasında değişir | |
| Jakobea | Jakoba, Reclinomonas | Serbest yaşayan, bazen çok gen açısından zengin mitokondriyal genomlara sahip loricate kamçılılar | |
| Metamonada veya POD | Preaksostyla | Oksimonadlar, Trimastix | Serbest yaşayan (Trimastix, Paratrimastix ) veya böceklerin arka bağırsaklarında yaşayan amitokondriat kamçılılar |
| Fornicata | Giardia, Carpediemonas | Amitokondriat, çoğunlukla ortak yaşamlar ve hayvanların parazitleri. | |
| Parabasalya | Trichomonas | Amitokondriat flagellatlar, genellikle böceklerin bağırsak kommensalleri. Bazı insan patojenleri. | |
| Neolouka | Malawimonadida | Malavimonas |
Soyoluş
[değiştir | kaynağı değiştir]
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||
Galeri
[değiştir | kaynağı değiştir]-
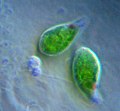
-
 Trypanosoma brucei (Euglenozoa: Kinetoplastida)
Trypanosoma brucei (Euglenozoa: Kinetoplastida) -
 Bodo sp. (Euglenozoa: Kinetoplastida)
Bodo sp. (Euglenozoa: Kinetoplastida) -
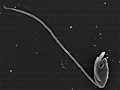 Percolomonas sp. (Percolozoa)
Percolomonas sp. (Percolozoa) -
 Stephanopogon sp. (Percolozoa)
Stephanopogon sp. (Percolozoa) -
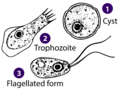
-
 Acrasis rosea (Percolozoa: Heterolobosea)
Acrasis rosea (Percolozoa: Heterolobosea) -
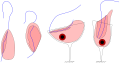 Jakobids (Jakobida)
Jakobids (Jakobida) -
-

-
 Giardia sp. (Metamonada: Fornicata: Diplomonadida)
Giardia sp. (Metamonada: Fornicata: Diplomonadida)











Kaynakça
[değiştir | kaynağı değiştir]- ^ Hampl (2009). "Phylogenomic analyses support the monophyly of Excavata and resolve relationships among eukaryotic "supergroups"". Proceedings of the National Academy of Sciences. 106 (10): 3859-3864. doi:10.1073/pnas.0807880106. PMC 2656170 $2. PMID 19237557.
- ^ "Phylogenomic analyses support the monophyly of Excavata and resolve relationships among eukaryotic "supergroups"". Proc. Natl. Acad. Sci. U.S.A. 106 (10): 3859-64. 2009. doi:10.1073/pnas.0807880106. PMC 2656170 $2. PMID 19237557.
- ^ Simpson, Ag (2006). "Comprehensive multigene phylogenies of excavate protists reveal the evolutionary positions of "primitive" eukaryotes". Molecular Biology and Evolution. 23 (3): 615-25. doi:10.1093/molbev/msj068. PMID 16308337.
- ^ a b Simpson (Dec 1999). "The ultrastructure of Carpediemonas membranifera (Eukaryota) with reference to the 'excavate hypothesis'". European Journal of Protistology. 35 (4): 353-370. doi:10.1016/S0932-4739(99)80044-3.
- ^ a b c "Cytoskeletal organization, phylogenetic affinities and systematics in the contentious taxon Excavata (Eukaryota)". International Journal of Systematic and Evolutionary Microbiology. 53 (6): 1759-1777. 1 Kasım 2003. doi:10.1099/ijs.0.02578-0. PMID 14657103. Birden fazla yazar-name-list parameters kullanıldı (yardım); Yazar
|ad1=eksik|soyadı1=(yardım) - ^ a b The Ancestor's Tale. 2016. ISBN 978-0544859937.
- ^ a b c Cavalier-Smith (2002). "The phagotrophic origin of eukaryotes and phylogenetic classification of Protozoa". International Journal of Systematic and Evolutionary Microbiology. 52 (2): 297-354. doi:10.1099/00207713-52-2-297. PMID 11931142.
- ^ Dawson (2013). "Alternative cytoskeletal landscapes: cytoskeletal novelty and evolution in basal excavate protists". Current Opinion in Cell Biology. 25 (1): 134-141. doi:10.1016/j.ceb.2012.11.005. PMC 4927265 $2. PMID 23312067.
- ^ "Revisions to the Classification, Nomenclature, and Diversity of Eukaryotes". The Journal of Eukaryotic Microbiology. 66 (1): 4-119. January 2019. doi:10.1111/jeu.12691. PMC 6492006 $2. PMID 30257078.
- ^ Brown (1 Şubat 2018). "Phylogenomics Places Orphan Protistan Lineages in a Novel Eukaryotic Super-Group". Genome Biology and Evolution. 10 (2): 427-433. doi:10.1093/gbe/evy014. PMC 5793813 $2. PMID 29360967.
- ^ Heiss (4 Nisan 2018). "Combined morphological and phylogenomic re-examination of malawimonads, a critical taxon for inferring the evolutionary history of eukaryotes". Royal Society Open Science. 5 (4): 171707. doi:10.1098/rsos.171707. PMC 5936906 $2. PMID 29765641.
- ^ Keeling (19 Ağustos 2019). "Progress towards the Tree of Eukaryotes". Current Biology. 29 (16): R808-R817. doi:10.1016/j.cub.2019.07.031. PMID 31430481.
- ^ Laura Wegener Parfrey; Erika Barbero; Elyse Lasser; Micah Dunthorn; Debashish Bhattacharya; David J. Patterson; Laura A. Katz (Aralık 2006), "Evaluating support for the current classification of eukaryotic diversity", PLOS Genetik, 2 (12), doi:10.1371/JOURNAL.PGEN.0020220, PMC 1713255 $2, PMID 17194223, Wikidata Q21090155
- ^ Tice (2021). "PhyloFisher: A phylogenomic package for resolving eukaryotic relationships". PLOS Biology. 19 (8): e3001365. doi:10.1371/journal.pbio.3001365. PMC 8345874 $2. PMID 34358228.
- ^ Kaynak hatası: Geçersiz
<ref>etiketi;Eme 2023isimli refler için metin sağlanmadı (Bkz: Kaynak gösterme)
Dış bağlantılar
[değiştir | kaynağı değiştir]- Açık Hayat Ağacı 6 Nisan 2023 tarihinde Wayback Machine sitesinde arşivlendi.
- taksonomikon 6 Şubat 2016 tarihinde Wayback Machine sitesinde arşivlendi.
- Hayat Ağacı Ökaryotlar 29 Ocak 2012 tarihinde Wayback Machine sitesinde arşivlendi.
- Hayat Ağacı: Jakobida 7 Şubat 2021 tarihinde Wayback Machine sitesinde arşivlendi.
- Hayat Ağacı: Fornicata 7 Şubat 2021 tarihinde Wayback Machine sitesinde arşivlendi.